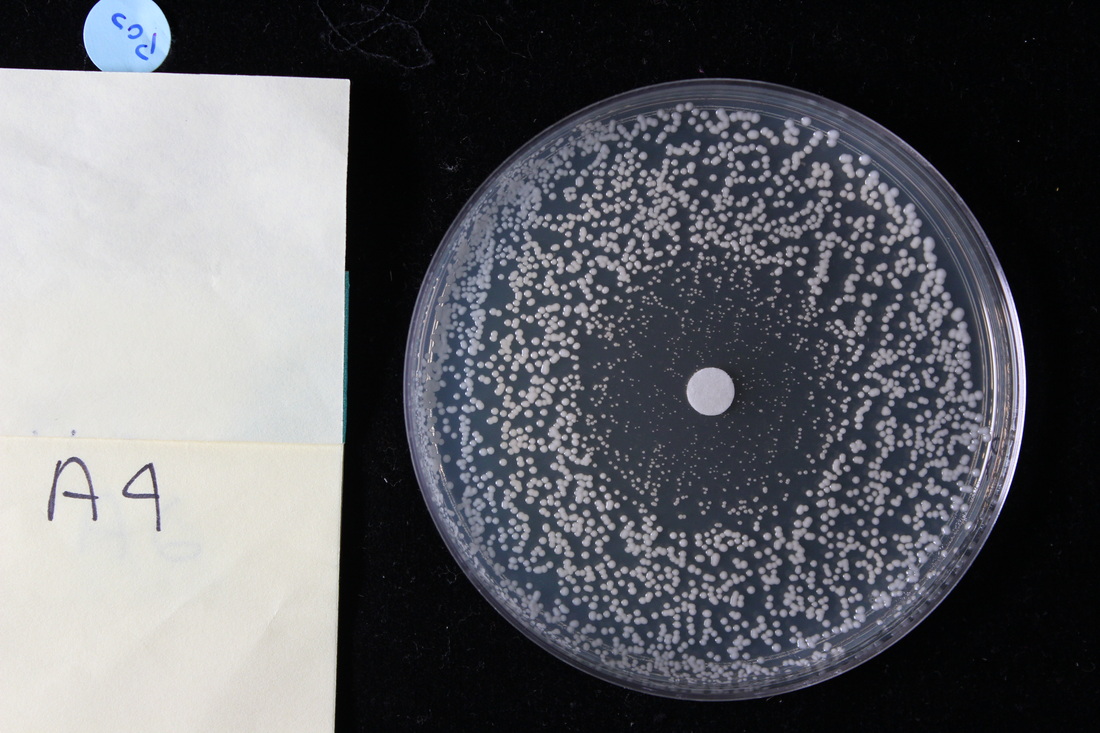
The acquisition of drug resistance
We examine the genotypic and phenotypic basis of drug resistance in fungal microbes. Current experiments in the lab involve the azole drug posaconazole, and the broad-spectrum antimicrobial boric acid. We have developed multiple new computational methods to quantify phenotypes including the quantification of drug tolerance from disk diffusion assays (the ability of a subpopulation of cells to grow slowly in high levels of drug) and biofilm formation rate from timelapse microscopy.
Some related manuscripts:
Gerstein and Berman 2020 Candida albicans genetic background influences mean and heterogeneity of drug responses
and genome stability during evolution to fluconazole. mSphere.
Ono, Gerstein and Otto 2017 Widespread genetic incompatibilities between first-step mutations during parallel
adaptation of Saccharomyces cerevisiae to a common environment. PLoS Biology.
Gerstein, Rosenberg, Hecht and Berman 2016 diskiImageR: quantification of resistance and tolerance to antimicrobial
drugs using disk diffusion assays. Microbiology
We examine the genotypic and phenotypic basis of drug resistance in fungal microbes. Current experiments in the lab involve the azole drug posaconazole, and the broad-spectrum antimicrobial boric acid. We have developed multiple new computational methods to quantify phenotypes including the quantification of drug tolerance from disk diffusion assays (the ability of a subpopulation of cells to grow slowly in high levels of drug) and biofilm formation rate from timelapse microscopy.
Some related manuscripts:
Gerstein and Berman 2020 Candida albicans genetic background influences mean and heterogeneity of drug responses
and genome stability during evolution to fluconazole. mSphere.
Ono, Gerstein and Otto 2017 Widespread genetic incompatibilities between first-step mutations during parallel
adaptation of Saccharomyces cerevisiae to a common environment. PLoS Biology.
Gerstein, Rosenberg, Hecht and Berman 2016 diskiImageR: quantification of resistance and tolerance to antimicrobial
drugs using disk diffusion assays. Microbiology

Predictability of evolution
We are interested in understanding the factors that promote or constrain parallelism in the evolutionary process. With collaborators we developed a statistical method to quantify genetic parallelism and are now applying this model to a meta-analysis of evolve-and-resequence experiments. We are conducting new experiments using in vitro evolution (evolution in the lab at large population sizes) and mutation accumulation (evolution at very low population sizes) to determine how differences in the environment and strain background influence the distribution of mutations and how often replicate lines evolve similarly or different.
Some related manuscripts:
Yeaman, Gerstein, Hodgins, Whitlock 2018 Quantifying how constraints limit the diversity of viable routes to adaptation.
PLoS Genetics.
Gerstein, Lo and Otto 2012 Parallel genetic changes and non-parallel gene-environment interactions underlie
nystatin resistance in yeast. GENETICS.
We are interested in understanding the factors that promote or constrain parallelism in the evolutionary process. With collaborators we developed a statistical method to quantify genetic parallelism and are now applying this model to a meta-analysis of evolve-and-resequence experiments. We are conducting new experiments using in vitro evolution (evolution in the lab at large population sizes) and mutation accumulation (evolution at very low population sizes) to determine how differences in the environment and strain background influence the distribution of mutations and how often replicate lines evolve similarly or different.
Some related manuscripts:
Yeaman, Gerstein, Hodgins, Whitlock 2018 Quantifying how constraints limit the diversity of viable routes to adaptation.
PLoS Genetics.
Gerstein, Lo and Otto 2012 Parallel genetic changes and non-parallel gene-environment interactions underlie
nystatin resistance in yeast. GENETICS.
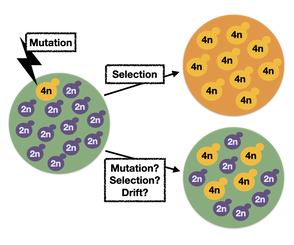
Rapid ploidy transitions during evolution
Ploidy variation (variation in the number of homologous chromosomes carried by an organism) is pervasive throughout the tree of life, even among closely related taxa. It remains largely unknown, however, when and why ploidy transitions arise and how they are maintained. Ploidy variation is widely seem in many contexts of fungal microbes, including in the natural environment, in clinical, environmental and industrial isolates, and rapidly arises under drug stress.
Some related manuscripts:
Gerstein and Sharp 2021 The population genetics of ploidy change in unicellular fungi. FEMS Microbiology Reviews.
Gerstein, Lim, Berman and Hickman 2017 Ploidy tug-of-war: Evolutionary and genetic environments influence the
rate of ploidy drive in a human fungal pathogen. Evolution
Gerstein, Chun, Grant and Otto 2006. Genomic convergence toward diploidy in Saccharomyces cerevisiae. PLoS Genetics
Ploidy variation (variation in the number of homologous chromosomes carried by an organism) is pervasive throughout the tree of life, even among closely related taxa. It remains largely unknown, however, when and why ploidy transitions arise and how they are maintained. Ploidy variation is widely seem in many contexts of fungal microbes, including in the natural environment, in clinical, environmental and industrial isolates, and rapidly arises under drug stress.
Some related manuscripts:
Gerstein and Sharp 2021 The population genetics of ploidy change in unicellular fungi. FEMS Microbiology Reviews.
Gerstein, Lim, Berman and Hickman 2017 Ploidy tug-of-war: Evolutionary and genetic environments influence the
rate of ploidy drive in a human fungal pathogen. Evolution
Gerstein, Chun, Grant and Otto 2006. Genomic convergence toward diploidy in Saccharomyces cerevisiae. PLoS Genetics
Recurrent vulvovaginal candidiasis
We are very excited to be beginning a new research program on vulvovaginal candidiasis (yeast infections). In collaboration with a local OB/GYN, we are acquiring yeast isolates from women with recurrent infections to examine the vaginal yeast and bacterial communities. We have been enrolling participants to study their isolates since summer 2022, with many projects planned.
Methods
In our lab we pair in vitro studies such as experimental evolution and high throughput phenotyping with bioinformatics and computational statistics to peer under the hood of evolution and understand the factors that underlie and drive evolution ot the genetic level. We have a particular interest in developing new quantitative methods to measure our parameters of interest in an unbiased manner. We are powered primarily by R. Current projects include a quantitative method to analyze biofilm formation, and the development of a bioconductor package to measure ploidy from flow cytometry of mixed cell populations.
We are very excited to be beginning a new research program on vulvovaginal candidiasis (yeast infections). In collaboration with a local OB/GYN, we are acquiring yeast isolates from women with recurrent infections to examine the vaginal yeast and bacterial communities. We have been enrolling participants to study their isolates since summer 2022, with many projects planned.
Methods
In our lab we pair in vitro studies such as experimental evolution and high throughput phenotyping with bioinformatics and computational statistics to peer under the hood of evolution and understand the factors that underlie and drive evolution ot the genetic level. We have a particular interest in developing new quantitative methods to measure our parameters of interest in an unbiased manner. We are powered primarily by R. Current projects include a quantitative method to analyze biofilm formation, and the development of a bioconductor package to measure ploidy from flow cytometry of mixed cell populations.